


 С учетом последних достижений биологии и медицины наиболее остро встает вопрос о времени начала человеческой жизни. От ответа на него зависит формирование отношения к проблемам абортов и контрацептивов, допустимости экспериментов над эмбрионами и вспомогательных репродуктивных технологий.
С учетом последних достижений биологии и медицины наиболее остро встает вопрос о времени начала человеческой жизни. От ответа на него зависит формирование отношения к проблемам абортов и контрацептивов, допустимости экспериментов над эмбрионами и вспомогательных репродуктивных технологий.
В работе проводится изучение эмбриологических событий развития эмбриона с момента оплодотворения до стадии нескольких первых дней. На основании анализа естественно-научных данных показывается, что жизнь человека как самостоятельного организма начинается с момента оплодотворения.
Введение
Последние несколько десятков лет в медицине отмечается значительное развитие репродуктивных технологий. В настоящий момент бесплодие преодолевается даже в тех случаях, когда половые клетки мужа не могут двигаться, а яйцеклетки женщины разрушены предшествующей химиотерапией.
Такие медицинские достижения ставят как перед учеными, так и перед обычными практикующими специалистами вопрос: когда начинается жизнь человека? В зависимости от ответа на него будут по-разному решаться важные биоэтические проблемы, связанные с лишением эмбриона жизни.
Православная биоэтика считает, что жизнь человека начинается с момента оплодотворения. Это положение оспаривается учеными на основании естественно-научных данных. Попытки опровергнуть их мнение на основании богословия или антропологии имеют мало успеха. Для ведения дискуссии с исследователями, обосновывающими время начала человека не с момента оплодотворения, необходимо использовать естественно-научные аргументы.
Попытки определения времени начала человеческой жизни в естественно-научном знании
Для современной науки характерны неопределенность и разногласие при определении временной границы, которая разграничивала бы эмбрион как биологический объект и как объект этический, к которому должно быть применимо отношение как к равноправному субъекту.
Некоторые признают достаточно поздние критерии появления человечности в эмбрионе. Так, Golderung утверждает, что о существовании человека можно говорить только после формирования нервной системы и начала мозговой деятельности. Однако экспертные комиссии различных государств, созданные для строгой регламентации исследований на эмбриональном материале, остановились на другой временной границе, разделяющей человеческий эмбрион от скопления неспециализированных клеток – 14 дней. Таким образом, в науке для этого срока закрепился термин «преэмбрион», выражающий усиленное биологическое значение этого объекта.
Тем не менее некоторые ученые признают более раннюю границу возникновения человечности. Так, в этом качестве рассматривается 8-й день развития. Высказываются мнения, что новый человеческий организм появляется приблизительно на третьи сутки развития после формирования поздней 8-клеточной стадии, в первый день развития на стадии 1 клетки после слияния генетического материала гамет (сингамии), непосредственно после слияния сперматозоида и яйцеклетки.
Возникают две принципиальные точки зрения: человек появляется сразу после слияния гамет или несколько позже (15 часов, 3 дня, 8 дней, 14 дней, 8 недель)?
Сущность вопроса о начале человеческой жизни
При оплодотворении гаметы сливаются, образуя зиготу. Гаметы являются составными частями родительских организмов. В гаметах происходят процессы жизнедеятельности. В зиготе также продолжается жизнь. Это продолжение жизни в зиготе приводит к рождению самостоятельного организма – ребенка. Значит, на этапе от оплодотворения до рождения происходит переход от жизненных процессов родительских организмов к жизненным процессам индивидуального организма. Итак, где же граница между жизнью половых клеток как составных частей взрослого организма и жизнью нового индивидуального организма?
Для ответа на этот вопрос необходимо определить, какие существуют критерии отдельного организма. То есть на основании чего мы можем утверждать, что тот или иной организм является самостоятельным, а не представляет собой колониальный многоклеточный организм или же часть материнского организма?
Для решения этой задачи нельзя ограничиться простым определением индивидуума. Индивидуум (индивид) (от лат. individuum – неделимое; особь) – особь, каждый самостоятельно существующий организм. По-другому можно сказать, что индивидуум – неделимое физическое целое, представляющее единичную сущность. С точки зрения наблюдателя, индивидуум – это то, что существует отдельно, самостоятельно. Это понятие, тем не менее, не всегда позволяет определить ту или иную форму жизни как самостоятельный организм. Например, Португальский кораблик (Physalia physalis) выглядит как самостоятельный организм, подходит под определение индивидуума, однако представляет собой колонию разнородных индивидуумов, каждый из которых имеет собственную нейронную сеть.
Человеческий организм в первые дни своего развития тоже представляет собой собрание нескольких клеток. Возникает вопрос: в это время представляет ли человек, подобно Physalia physalis, колонию клеток-организмов, плохо прикрепленных друг ко другу и слабо взаимодействующих между собой?
Очевидно, чтобы ответить на вопросы, поставленные выше, необходимо использовать иные понятия – те, которые помогают определить, что же такое самостоятельный организм. Одним из таких понятий может стать понятие «оператор», которое использует Gerard A. J. M. (Jagers op Akkerhuis (2010) в своей теории иерархии операторов.
Теория иерархии операторов
Теория иерархии операторов возникает в ответ на поиски определения жизни, организма, смерти. Gerard исследует уровни организации материи. Он последовательно анализирует усложнение материи в адронах, атомах, молекулах, прокариотической и эукариотической клетках, многоклеточном организме. Для такого анализа вводится понятие оператора (operator). Оператор – обладающая автономной активностью сущность, которая действует в окружающей среде без потери ее индивидуальной организации. Для живого оператора необходимо формирование функциональной и структурной замкнутости. В живых системах функциональную замкнутость представляют собой циклы автокаталитических реакций, структурной же замкнутостью является клеточная мембрана. Клеточная мембрана и цикл каталитических реакций позволяют клетке поддерживать ее морфологическую структуру и функционировать как единой целое, действующее в окружающей среде без потери ее индивидуальной организации, делая её таким образом простейшим живым оператором.
Многоклеточный организм в системе операторов может быть определен следующим образом: это группа взаимно прикрепленных клеток с обязательным возвратным взаимодействием, функционирующая как единое целое. Таким образом, мы можем рассматривать группу клеток как многоклеточный организм, если между клетками формируются а) структурные и б) функциональные замкнутости.
Для определения типа связи между клетками, необходимого для признания многоклеточного организма единым, Gerard рассматривает эволюционный процесс формирования многоклеточности. Gerard учитывает, что позвоночные животные в процессе своего онтогенеза повторяют этапы развития многоклеточного организма, поэтому приходит к выводу, что во время раннего своего развития они представляют собой колонию. Единым многоклеточным организмом эмбрион становится только после возникновения щелевидных контактов между клетками, а до этого организм может рассматриваться лишь в качестве колонии.
Тем не менее стоит предполагать, что не только щелевидные контакты могут формировать структурную замкнутость и, следовательно, могут обусловливать отнесение того или иного организма к многоклеточному, а не к колонии. Структуры, отграничивающие живой объект от окружающей среды и поддерживающие его индивидуальную организацию, являются для организма своеобразной структурной замкнутостью.
Функциональная замкнутость требует взаимообратной связи, то есть взаимной автокаталитической зависимости клеток. В процессе эмбрионального развития организма такую функциональную возвратную взаимосвязь как необходимый элемент многоклеточного организма обнаруживают процессы дифференцировки, которые происходят при взаимодействии клеток. Данное явление называется индукцией. Индукция – процесс, посредством которого одна область зародыша, взаимодействуя с другой, побуждает эту область развиваться в направлении ином, чем она развивалась бы без этого воздействия.
Эксперименты Г. Шпемана и Х. Мангольд по пересадке зачатка будущей спинной стороны (дорсальной губы бластопора) показали, что эта область является «организатором», способным «инструктировать <запускать – прим. свящ. Р. Т.> формирование новых эмбриональных осей». Таким образом, было доказано индуктивное влияние клеток данного зачатка (дорсальной губы) на окружающие клетки и изменение направления их развития. Однако и сами клетки дорсальной губы становятся способными проявлять свойства первичной эмбриональной индукции только под воздействием дорсальных вегетативных клеток бластулы, названных центром Ньюкупа. Таким образом, в многоклеточном организме наблюдается серия последовательных влияний клеток друг на друга в процессе дифференцировки.
Индуктивные влияния организатора проявляются и при процессах регенерации утраченных частей тела организма. Так, эксперименты Э. Браун с пересадкой различных участков гидры показали, что в гидре имеется участок тела, выполняющий функцию организатора. Таким образом, в гидре как многоклеточном организме существует тесная взаимосвязь клеток, проявляющаяся во взаимном влиянии одних клеток на другие и способности вторых клеток воспринимать эти влияния.
В колониальном организме такого взаимного влияния клеток друг на друга нет (следовательно, нет и функциональной замкнутости). Губки могут образовывать колониальный организм, в котором клетки дифференцированы и имеют тенденцию к образованию тканей. При этом регенерация, видимо, связана с бесполым размножением организмов, составляющих колонию. Значит, у губок отсутствует единый организующий центр, контролирующий репаративные процессы индукционным влиянием, отсутствуют и части, воспринимающие индукцию. А следовательно, отсутствует функциональная замкнутость, необходимая для формирования многоклеточного организма.
Таким образом, индукционное влияние клеток в процессе их дифференцировки, в том числе во время эмбрионального развития, отражает их тесную функциональную взаимосвязь и объединяет их в единый многоклеточный организм. Рассмотрим, на какой из стадий происходит формирование структурной и функциональной замкнутостей, необходимых для признания эмбриона самостоятельным организмом.
Применение теории операторов к многоклеточному эмбриону
Поздняя 8-клеточная стадия
В качестве необходимой структурной замкнутости в теории операторов признаются щелевидные контакты. Они формируются при компактизации бластомеров, что наблюдается на поздней 8-клеточной стадии.
Функциональная замкнутость (строгое взаимодействие и обратная взаимосвязь клеток многоклеточного организма) четко видна на стадии 16-клеточной морулы. В это время происходит достаточно точная фиксация дальнейшего пути развития каждого из бластомеров: из наружных клеток формируется поверхностный слой клеток – трофобласт (впоследствии потомки этих клеток выполняют питательную функцию и не участвуют в построении непосредственно зародыша человека; они войдут в состав плаценты), из внутренних – внутриклеточная масса (из них в дальнейшем будут формироваться ткани плода). При этом «к внутренним клеткам 16-клеточного зародыша иногда добавляется одна наружная клетка при переходе к 32-клеточной стадии». «К стадии 64 клеток внутриклеточная масса и клетки трофобласта превращаются в полностью сформированные клеточные слои, ни один из которых не поставляет клеток другой группе». Данные события показывают строгое взаимодействие клеток друг с другом в процессе их дифференциации.
Наличие такого взаимодействия (функциональной замкнутости) и щелевидных контактов (структурной замкнутости) согласно теории операторов доказывает формирование самостоятельного организма на поздней 8-клеточной стадии.
Ранняя 8-клеточная и 4-клеточная стадия
Указанные выше процессы взаимодействия бластомеров происходят и на стадиях более ранних, чем поздняя 8-клеточная, определяя своеобразную функциональную замкнутость в едином многоклеточном организме.
Еще до компактизации бластомеров на поздней 8-клеточной стадии в цитоплазме бластомеров происходит процесс поляризации плазматической мембраны, когда имеет место ее перестройка. Gilbert приводит исследования, показывающие, что при мечении некоторых поверхностных молекул мембран 4-клеточных бластомеров красители беспорядочно распределяются по поверхности, а на стадии незавершенного 3-го деления они обнаруживаются на полюсах, наиболее удаленных от центра бластулы. Важно, что данный процесс происходит до начала компактизации и при обязательном контакте бластомеров. Gilbert заключает, что данное перемещение является, видимо, результатом межклеточных взаимодействий, так как оно выявляется при выделении из зародыша двух связанных между собой бластомеров, но не бластомеров, изолированных поодиночке. Учитывая такое взаимодействие бластомеров, стоит предположить, что отсутствующие на этой стадии плотные щелевидные контакты не препятствуют 4-клеточному эмбриону быть самостоятельным организмом.
Структурную замкнутость, отграничивающую организм от окружающей среды и позволяющую ему существовать в качестве индивидуальной сущности, на этой стадии обеспечивает блестящая оболочка. Вылупление эмбриона из блестящей оболочки, то есть хэтчинг, происходит приблизительно на 5-й день развития на стадии бластоцисты. В это время между бластомерами уже имеются достаточно плотные межклеточные соединения. Количество бластомеров достигает более 100 клеток. До хэтчинга блестящая оболочка помогает рыхло прикрепленным друг ко другу бластомерам удерживаться вместе и тем самым представляет ту самую структурную замкнутость, необходимую для признания живого объекта самостоятельным организмом.
4-клеточная и 2-клеточная стадии
Функциональная взаимосвязь между бластомерами 4-х и даже 2-клеточного эмбриона может быть показана на примере дифференцировки бластомеров на этих стадиях. Вопрос о дифференцировке бластомеров на таких ранних стадиях возникает в связи с исследованием пред-моделей поведения бластомеров, то есть условий, которые в последующем обусловливают выбор бластомерами определенного пути развития: в клетки внутриклеточной массы (а следовательно, в клетки эмбриона) или же в клетки трофобласта (а значит, во внеэмбриональные ткани).
Как уже указывалось выше, положение клеток в 16-клеточном эмбрионе определяет ее судьбу: наружные клетки образуют трофобласт, внутренние – внутриклеточную массу. Однако существуют ли более ранние факторы, определяющие судьбоносный выбор бластомеров, или же их случайное положение внутри 16-клеточного зародыша диктует направление их развития?
В работе Takaoka K. (2014) приводится гипотеза, основанная на изучении эпигенетических факторов различия генетического материала, полученного от материнской и от отцовской особей мыши. Во-первых, автор сообщает, что эпигенетические различия уже имеются в мужской и женской половинах генома на стадии пронуклеусов. После оплодотворения в спермальной ДНК белки замещаются ацетилированными гистонами, а не метилированными. В то время как ДНК и ассоциированные с нею гистоны (например, тип гистонов Н3) женского пронуклеуса высоко метилированы. Данные изменения сохраняются на 2-клеточной стадии. А на 4-клеточной стадии было показано, что эпигенетические различия бластомеров влияли на дальнейшую судьбу бластомеров. Повышенный уровень метилированных гистонов Н3 предпочтительно обусловливали дифференциацию бластомеров во внутриклеточную массу. При этом введение в один из 2-клеточных бластомеров фермента, который способствует переносу метильных групп на гистоны (в том числе и на Н3), обуславливало преимущественную дифференциацию этого бластомера во внутриклеточную массу. Во-вторых, асимметрия родительских пронуклеусов увеличивается за счет уменьшению количества метильных групп (=деметиляция) на самой ДНК. Так, ДНК мужского пронуклеуса активно деметилируется при посредничестве диоксигеназы. В-третьих, женский пронуклеус защищен от действия этого фермента посредством фактора, связанного с одним из метилированных гистонов, который есть в женском пронуклеусе, но отсутствует в мужском. Поэтому предполагают, что метиляция ДНК и метиляция гистонов имеют взаимное влияние и могут регулировать друг друга.
На основании изложенного была сформирована гипотеза, согласно которой существующая асимметрия (метиляция родительских ДНК или гистонов) влияет на дальнейшую дифференцировку бластомеров (рисунок 1).
При условии, что зиготическая транскрипция гипотетического фактора начинается на стадии 2 клеток, клетки с материнской модифицированной ДНК (красные точки) на 2-х и 4-клеточной стадии экспрессируют неизвестные гены (зеленый цвет цитоплазмы), которые будут влиять на положение бластомеров (внутреннее или наружное) и их последующее судьбоносное решение стать клетками внутриклеточной массы (ICM) или трофобласта (TE) на более поздней стадии. Вторая клетка (на рисунке 2: в 4-клеточной стадии вторая сверху) содержит более низкий продукт этого гена (светло-зеленая цитоплазма), который наследуется из предшествующей клетки 2-клеточной стадии. Однако это может оказаться достаточным для экспрессии регуляторных генов, которые определяют дифференциацию этого бластомера во внутриклеточную массу (ICM).
Таким образом, высказанная гипотеза показывает возможную причину, по которой происходит разная дифференциация рядом расположенных бластомеров на стадии 4-х и даже 2-клеточного зародыша.
Следует заметить, что использование бластомерами того или иного пути дифференциации возможно только в присутствии других бластомеров. Толькопри контакте и влиянии друг на друга будет реализован тот вектор развития в клетки внутриклеточной массы или клетки трофобласта, который, согласно гипотезе, может быть предположен уже на 2-клеточной стадии.
Таким образом, можно говорить, что данный процесс дифференцировки на стадии двух бластомеров свидетельствует о формировании функциональной замкнутости, то есть возвратного взаимодействия и взаимной автокаталитической зависимости 2 клеток.
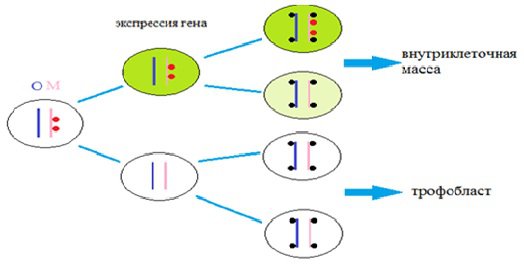
Рисунок 1. Гипотетическая модель возникновения условий, определяющих образец поведения ранних бластомеров, основанная на эпигенетической асимметрии родительских геномов.
Показана модель возникновения условий, определяющих поведение ранних бластомеров, основанная на эпигенетической асимметрии родительских геномов. Материнский геном обозначен розовым, отцовский – синим цветом. Асимметричные модификации родительских геномов (метиляция ДНК или гистонов) изображены красными точками на материнской хромосоме. В этой модели асимметричные модификации не наследуются дочерними хромосомами после клеточного деления. Новые модификации (изображены черными точками) случаются после 2-го клеточного деления равным образом на материнских и на отцовских по происхождению хромосомах.
Учитывая, что в человеческом эмбрионе на рассматриваемых стадиях имеется замкнутость как структурная (Zonapellucida), так и функциональная (начавшиеся процессы дифференцировки бластомеров, которые заключаются в избирательном метилировании гистонов или ДНК и влияют на дальнейшую дифференцировку бластомеров при обязательном их взаимном контакте), можно говорить о формировании оператора, а следовательно, и самостоятельного организма уже на стадии 2 клеточного зародыша.
Позиция Gerard по отношению к многоклеточному эмбриону и ее опровержение
Согласно Gerard, мы можем рассматривать живую систему как многоклеточный организм, если между клетками формируются структурные и функциональные замкнутости. Gerard говорит, что у позвоночных организмов структурная замкнутость образуется только после возникновения щелевидных контактов между клетками, а до этого организм может рассматриваться лишь в качестве колонии. Указанные щелевидные контакты формируются при компактизации бластомеров, что наблюдается на поздней 8-клеточной стадии. Таким образом, следуя рассуждениям автора, вплоть до ранней 8-клеточной стадии включительно мы не можем говорить о появлении индивидуального организма – человека. Именно этим автор обосновывает возможность появления двух монозиготных близнецов из одного многоклеточного эмбриона.
Действительно, бластомеры на 4-клеточной стадии не имеют плотных контактов между собой, располагаются рыхло и между ними имеются большие пространства. Мнение Gerard подтверждается исследованием (HildeVandeVelde, 2008), в котором было произведено разделение шести 4-клеточных эмбрионов. Полученные 24 бластомера культивировались до стадии бластоцисты. В результате были получены 16 бластоцист хорошего качества с наличием внутренней клеточной массы и трофобласта. Однако появление двух монозиготных близнецов при разделении эмбриона может происходить и после 8-клеточной стадии, вплоть до начала гаструляции, то есть до 14-го дня, что ставит под сомнение выводы Gerard.
Кроме того, надо иметь в виду, что исследования VandeVelde происходили после искусственного лишения эмбриона блестящей оболочки. Разделенные отдельные бластомеры в дальнейшем переносились в блестящую оболочку, где культивировались до стадии бластоцисты. Получается, что каждый отдельный организм (в данном исследовании доведенный до стадии бластоцисты) должен был иметь контакт с блестящей оболочкой. Это в свою очередь подтверждает особую роль блестящей оболочки в формировании самостоятельного организма.
Таким образом, исследование по разделению 4-клеточной бластулы на бластомеры не подтверждает мнение Gerardо том, что до возникновения плотных контактов эмбрион представляет лишь колонию клеток, а не самостоятельный организм. Результаты VandeVelde как раз показывают, что функцию структурной замкнутости, необходимой для признания организма самостоятельным оператором, до формирования плотных межклеточных контактов в эмбрионе выполняет блестящая оболочка.
К тому же стоит заметить, что в этом исследовании после расщепления 4-клеточных эмбрионов на бластомеры последние только на 3-й день раздельной культивации (72 часа) совершали первое деление на 2 клетки. Учитывая, что первое деление зиготы обычно происходит приблизительно через 30 часов после оплодотворения (а последующие деления еще быстрее), стоит предполагать, что избыточное время (72 ч вместо 30 ч), необходимое бластомерам для начала деления, используется как раз для исправления наследуемой эпигенетической асимметрии бластомеров. Такое включение компенсаторных механизмов для начала повторного деления подтверждает наличие функциональной взаимозависимости (=замкнутости) ранних бластомеров.
Таким образом, данные явления не опровергают утверждения о наличии в 2-х и 4-клеточном эмбрионе структурной и функциональной замкнутостей. Следовательно, на данных этапах можно говорить об эмбрионе как самостоятельном организме.
Применение теории иерархии операторов для решения вопроса о времени начала человеческой жизни на стадии зиготы
Развитие неоплодотворенной яйцеклетки как части организма матери
После овуляции яйцеклетка, окруженная клетками кумулюса, попадает в маточные трубы. Если в течение 24 часов не происходит оплодотворение, то яйцеклетка претерпевает комплекс изменений, называемых старением яйцеклетки. Среди механизмов старения выделяют аномальное спонтанное повышение кальция. Это приводит к фрагментации яйцеклетки и другим механизмам апоптоза. Немалую роль в программируемой гибели яйцеклетки играет дисфункция митохондрий, наблюдаемая при ее старении. Причиной этого может стать повреждение митохондрий реактивными кислородсодержащими веществами (такими, как супероксид). Кроме того, супероксид индуцирует затвердение блестящей оболочки, разрушение цитоплазматических микротрубочек. Среди механизмов старения яйцеклеток выделяют эпигенетические изменения их ДНК. Все это приводит к структурным изменениям стареющего ооцита: нарушение структуры микроворсинок и их отпочковывание, затвердевание блестящей оболочки, миграцию кортикальных гранул и их частичный экзоцитоз, разрушение микрофиламентов, изменение веретена, нарушение расположения хромосом в метафазной пластинке, рассеивание и в некоторых случаях их деконденсация.
Таким образом, процессы, происходящие в яйцеклетке при старении, направлены на ее разрушение. Именно поэтому способность яйцеклетки к оплодотворению уменьшается с течением времени после овуляции и прекращается через 24 часа после нее.
Развитие оплодотворенной яйцеклетки
Зигота – клетка, которая образуется при слиянии яйцеклетки и сперматозоида. После слияния гамет происходит каскад реакций, защищающий яйцеклетку от проникновения остальных спермиев. В первую очередь происходит быстрый блок полиспермии путем изменения электрического потенциала плазматической мембраны яйцеклетки [14]. Он сохраняется недолго. К моменту его окончания формируется медленный блок полиспермии путем экзоцитоза кортикальных гранул, которые содержат ферменты, модифицирующие рецепторы прозрачной оболочки. Сперматозоиды теряют способность связываться с ними.
Повышение внутриклеточного содержания ионов кальция активирует белковый синтез и синтез ДНК. Увеличение потребления оплодотворенной клеткой кислорода (у некоторых организмов) наблюдается уже через 1 мин после слияния гамет в то время, как активация синтеза белка – только через 5-10 мин .
После слияния гамет их ядра формируют пронуклеусы. В каждом из них происходит синтез ДНК. Данный процесс продолжается в течение 8-10 часов. В это время они направляются навстречу друг к другу. Их встреча происходит у человека приблизительно через 15 часов после слияния гамет. Однако настоящего слияния ядер (сингамии) в человеческой зиготе не происходит. При соединении пронуклеусов происходит разрушение их оболочек, хроматин конденсируется в хромосомы, которые располагаются по-отдельности на общем митотическом веретене деления. Общее ядро формируется только на 2-клеточной стадии.
Имеются работы, которые показывают начало транскрипции, то есть проявления активности собственного генома эмбриона, уже на стадии одной клетки. Так, Martin-McCaffrey (2004) в своем исследовании определил у мышей белок RGS14, участвующий в построении веретена деления. Белок необходим для первого клеточного деления и при этом отсутствует в гаметах, в которых отсутствует также и его м-РНК. Авторы сделали вывод, что он вырабатывается denovo на стадии 1-клеточного эмбриона, являясь показателем активации генома эмбриона уже на стадии пронуклеусов.
Другое исследование, Daniels R. (1997), показало, что ген XIST, регулирующий инактивацию Х-хромосомы у человека, активизируется уже на 1-клеточной стадии эмбриона и его активность увеличивается к моменту 8-клеточной стадии.
Имеются и другие работы, показывающие начало транскрипции на 1-клеточной стадии. Развитие на стадии 2-клеточного зародыша обусловлено активностью транскрипции на предыдущей стадии.
Кроме активности генов на стадии пронуклеусов, отцовская и материнская половинки генома активно взаимодействуют: транскрипция материнской ДНК подавляется мужским пронуклеусом, транскрипция которого в 5 раз выше материнского .
Сравнение процессов, происходящих в оплодотворенной и неоплодотворенной яйцеклетках с позиции теории операторов
Итак, если яйцеклетка не оплодотворяется, то происходящие в ней процессы направлены на ее разрушение. Если же происходит оплодотворение, то начинается комплекс реакций, направленных на рост и развитие: синтез ДНК, экспрессия генов, взаимодействие пронуклеусов, усиление метаболизма, увеличение потребления кислорода. После оплодотворения появляется иная модель поведения клетки. Кроме того, важно отметить взаимодействие ооцита и клеток кумулюса до и после оплодотворения. Клетки кумулюса, будучи соединенными с ооцитом щелевидными контактами, играют чрезвычайную роль в созревании ооцита, движению его в маточные трубы до оплодотворения, участвуют в процессе оплодотворения (см. рисунок 2) . И если же оплодотворение не происходит, то клетки кумулюса продолжают влиять на ооцит, усиливая его старение . После же оплодотворения ооцит отграничивается от окружающих его клеток кумулюса посредством кортикальной реакции и затвердевания блестящей оболочки.
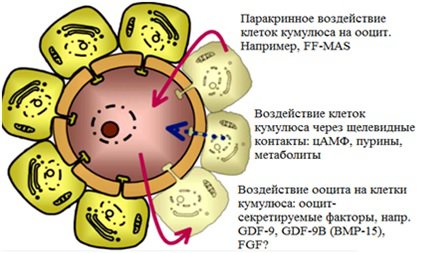
Рисунок 2. Взаимодействие неоплодотворенной яйцеклетки и клеток кумулюса
Учитывая тесную структурную взаимосвязь клеток кумулюса и неоплодотворенной яйцеклетки, их сильное функциональное взаимодействие, можно утверждать, что неоплодотворенная яйцеклетка представляет собой часть многоклеточного материнского организма.
После оплодотворения происходит разрушение структурной и функциональной взаимосвязей яйцеклетки и клеток кумулюса. Зигота отграничивается от клеток кумулюса при помощи кортикальной реакции и утолщения Z. Pellucida – формируется структурная замкнутость. Кроме того, в зиготе, как было отмечено выше, после оплодотворения начинается цепь автокаталитических реакций оплодотворенной яйцеклетки, поддерживающих и развивающих ее структуру, появляется иная модель поведения клетки: механизмы старения сменяются механизмами роста и развития, то есть формируется функциональная замкнутость, изолирующая зиготу от материнского организма.
Таким образом, формирование функциональной замкнутости, наряду с отграниченностью зиготы от окружающего мира (структурная замкнутость), позволяет говорить о появлении на стадии зиготы, то есть на стадии 1-клеточного эмбриона, самостоятельного организма, то есть индивидуума.
Выводы
Анализ процессов, происходящих после оплодотворения зиготы и на начальных этапах развития эмбриона, показывает, что самостоятельный организм, сохраняющий свою структуру, самостоятельно действующий в окружающей среде на основании уникального генетического материала, доставляемый в новый организм при помощи гамет, появляется уже на 1-клеточной стадии эмбрионального развития.
Данный генетический материал, задающий вектор развития нового организма, постепенно реализуется в виде качеств и свойств, но начинает проявляться уже на стадии пронуклеусов, то есть еще до объединения отцовской и материнской ДНК в едином ядре.
Изменение модели поведения после оплодотворения (старение неоплодотворенной яйцеклетки сменяется ростом и развитием оплодотворенной, усилением её метаболизма, отграничением от окружающего пространства) в течение нескольких секунд после оплодотворения подтверждает то, что самостоятельный организм появляется сразу после слияния сперматозоида и яйцеклетки.
В последующем своем развитии на многоклеточной стадии эмбрион также проявляет себя как единая сущность, обладающая структурной и функциональной замкнутостью. Это доказывает наличие в нем самостоятельного отдельного организма, а не колониальной формы жизни.
священник Тарабрин Роман











